L. J. S. 15 Feb 87
| TABLE OF CONTENTS (Click on Chapter titles to go to chapter) |
||
|---|---|---|
| Chapter Title | Page | |
| ABSTRACT | ii | |
| ACKNOWLEDGEMENTS | iii | |
| LIST OF FIGURES | v | |
| LIST OF TABLES | vii | |
| INTRODUCTION | 1 | |
| GEOMORPHOLOGY AND HYDROLOGY | 2 | |
| SOILS | 1 | |
| CLIMATE | 7 | |
| VEGETATION | 13 | |
| Government Land Office (GLO) Survey | 13 | |
| Floristic Reconstruction | 16 | |
| FAUNA | 26 | |
| Mammals | 26 | |
| Birds | 31 | |
| Amphibians and Reptiles | 34 | |
| Fishes | 38 | |
| Mollusks | 42 | |
| Other Invertebrates | 44 | |
| Conclusions | 45 | |
| CONCLUSIONS | 46 | |
| APPENDIX | 48 | |
| REFERENCES CITED | 51 | |
| LIST OF FIGURES (Click on Figure number to go to graphic) |
|
|---|---|
| Figure | |
| Figure 1. Moraines of the Upper Sangamon Basin. | |
| Figure 2. Climatograph Displaying Monthly Mean Temperature and Precipitation for Urbana. | |
| Figure 3. The illustrated faunal element is a fragment of the Left Inferior M3 9-15(?)/24 of Woolly Mammoth (Mammuthus primigenius) recovered from a residential subdivision development located less than a mile west of Parkland College, Champaign County. | |
| Figure 4. The Original Land Survey Map of Mahomet Township, Champaign County. | |
(Click on Table number to go to graphic) |
||
|---|---|---|
| Table | Page | |
| Table 1. | Calculations for the F-Statistic Testing for Significant Differences in the Mean Distances of the Three Most Common Species in the GLO Survey Moraines of the Upper Sangamon Basin. | 16 |
| Table 2. | Forest Species Identified in the GLO Survey. | 19 |
| Table 3. | Importance Values for the First Five Dominants of the Upland Forest in Five Studies of the Sangamon Drainage. | 21 |
| Table 4. | Importance Values of the Five Leading Dominants of the Floodplain Forest From Three Sites in the Sangamon Drainage. | 23 |
| Table 5. | Historically Reported Mammals, Preferred Habitats, and Presence in the Archaeological Record. | 28 |
| Table 6. | Birds Representative of Several Habitats Within the Park. | |
| Table 7. | Amphibians and Reptiles of the Park and Preferred Habitats. | 35 |
| Table 8. | Additional Reptile Species Recovered from the Pabst Site. | 37 |
| Table 9. | Fish Species Characteristic of Sangamon Drainage Habitats. | 39 |
| Table 10. | Mussels of the Sangamon River, Piatt County. | 43 |
| Table 11. | An Inventory of Terrestrial Snails from Two Prairie Groves in Champaign County. | 44 |
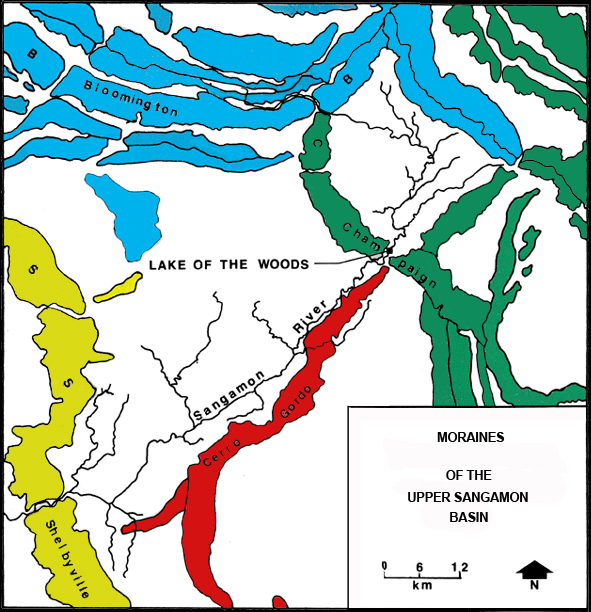
Woodfordian ice began to disrupt its headwaters. The final reconfiguration of
the headwaters began less than 20,000 years ago when the Woodfordian glacier
reached maximum at the Shelbyville Moraine. As the ice wasted to the north and
east, it reestablished a front marked by the Cerro Gordo Moraine. Over this
part of the Cerro Gordo's course it forms a relatively straight line from
Mahomet to Decatur. As the river passes through the Shelbyville Drift, its
east wall is defined by this moraine. How the river penetrated the Champaign
Moraine is an unresolved issue. David Reinertson of the Illinois Geological
Survey (personal communication) suggests that the gap in the moraine at Mahomet
indicates the presence of a subglacial channel issuing meltwater from beneath
the glacier's toe. As the rate of flow increased from the wasting Woodfordian
ice, the notch was more extensively eroded and assumed its present morphology.
These meltwaters also cut the valley to its present configuration in the Cerro
Gordo outwash (Cote et al. 1969:14). Above the Champaign Drift, the valley
extends roughly 10 miles on to the back slope of the Bloomington Moraine. This
condition was precipitated when the Peoria Sublobe of the Woodfordian
glaciation readvanced and overrode the northwestern extreme of the Champaign
Moraine between 14,000 and 15,000 years ago (Willman and Frye 1970:94). Miller
(1973:28) states that in the ensuing 14,000 years the river valley has remained
quite stable with slow rates of lateral migration and floodplain sedimentation
(0.21 mm/yr). Within the park the valley is never more than 400 m in width.
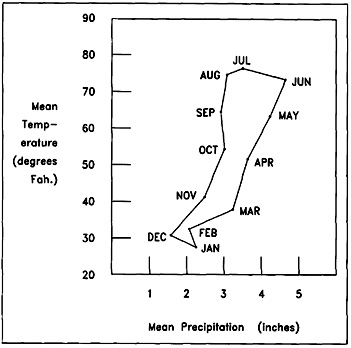
saw only minor climatic oscillations with the temperate, continental conditions
of today seemingly firmly stabilized.
Perhaps no discussion of Late Pleistocene mammalian assemblages would be complete without some reference to the Woolly Mammoth (Mammuthus primigenius). In the fall of 2009 a molar fragment recovered from the excavation of a waterline in a residential subdivision located less than a mile west of the college was brought to our attention. While all such recoveries are by definition suspect, I have little reason to doubt the veracity of the provenience. The find would have come from on or near the crest of the Pesotum Moraine (Willman et al 1975:234-235). The Pesotum is a recessional moraine associated with the Decatur Sub-lobe of the Woodfordian glaciation. While, owing to the item's apparent isolated condition, the possibility of glacial redeposit remains a possibility, the most likely interpretation is that of direct deposition and primary context. If such is the case then the mammoth would have been on this landscape sometime after 16,000 BP and prior to the vegetative succession from open spruce woodland/tundra at 13,800 BP.
Students in my fall 2009 Introduction to Archaeology (ANT 200) class determined that they would research and illustrate the molar (contributors included: Whitney Hoveln, Teri Jolley, Thuy H. Luu, Joseph A. Miebach, Kathryn R. Rumer, Brittany K. Salm, Kyle W. Washburn, and Matthew T. Wilson). We here present some of their effort.





Government Land Office Survey - The GLO survey provides a useful
reconnaissance of the park's vegetational matrix as it existed in 1822. In
this year, surveyor Richard T. Holliday, under the contract of Elias and Worton
Rector, completed the land survey of what is today known as Mahomet Township
(Township 20 North, Range 7 East), Champaign County (Holliday 1822). The
actual survey was completed between July 8 and 13. The surveyor was instructed
to establish section and quarter section points. Where possible, witness trees
were to be employed for relocating these points; otherwise a post was set in a
mound at the designated location. When witness trees were used, Holliday
generally recorded two for the section corners and one for the quarter
sections. Information about the witness trees that is entered in his field
notes includes species name, diameter at breast height, bearing from the
reference point, and its distance from the reference point. At the completion
of each mile Holliday typically provides a rather cryptic synopsis of the
vegetation, landforms, and soil. On prairie transects the vegetational
reference was simply "prairie" and no attempt was made at identifying species.
Floristic Reconstruction - Having earlier in the paper
demonstrated a series of paleoclimatic episodes, the following depiction of the
research area must be tempered with the knowledge that we are here describing a vegetational distribution and mix that best reflects the climate of the late Neoglacial. During the last 8,300 years, range shifts for the
native communities are certainly indicated by the climatic data. Moreover, on
the basis of the palynological data, the phytosociologies of the present
communities display an inadequate relationship to the flora present in this
locale prior to the emergence of prairie at 8,300 BP to be of much use in their
interpretation. Nonetheless, for the bulk of the prehistoric sequence, these
floristic elements would seem to form the foundation of the park's vegetation.
The comparison is based upon the five species from each setting having the
highest Importance Value (Importance Value = relative density + relative
dominance). The oaks clearly dominate each stand. The hickories are
surprisingly variable in their distribution, with shagbark ranked only ninth at
Hart Woods (IV = 2.1). In fact, perhaps the most striking result of this
comparison is the degree of variability we find in both species frequency and
diversity. The oak-hickory forest was apparently not particularly homogeneous,
and its floristic resources were not evenly distributed throughout its range.
Moreover, the observation by Root et al. (1971:36) that hickories were
reproducing more successfully than the oaks in Hart Woods implies temporal
variations in species frequency within a single site. These spatial and
temporal variations within the oak-hickory forest likely had implications for
prehistoric site selection and function.
In conclusion, the park's most expansive presettlement communities included
the Dry-mesic Upland Forest, the Wet-mesic Floodplain Forest, and the Dry-mesic
Savanna. The floristic resource potential of the Mesic Prairie, the two
aquatic communities, and Cultural Disturbance community was probably limited.
F. King (1984) provides an excellent analysis of the ethnobotanical
implications of the study area, and Jones and Bell (1974) offer an annotated
checklist of the vascular flora of the Sangamon catchment.
I. Mammals:
II. Birds:
III. Amphibians and Reptiles:
Table 7. Amphibians and Reptiles of the Park and Preferred Habitats.
Species - Common Name , Preferred Habitat1
Table 8. Additional Reptile Species Recovered from the Pabst
Site.1
IV. Fishes:
Table 9. Fish Species Characteristic of Sangamon Drainage
Habitats.1
V. Mollusks:
Table 10. Mussels of the Sangamon River, Piatt County.1
the species parameters that we might encounter within our study area. The
checklist includes a total of 27 species. The presence of both aquatic and
terrestrial snail shells from prehistoric sites in Illinois (Parmalee
1968:107) lends strong support for an interpretation of conscious exploitation.
Functions were likely both dietary and ornamental.
Table 11. An Inventory of Terrestrial Snails from Two Prairie Groves in
Champaign County.1
1 - Adapted from Zetek (1918).
VI. Other Invertebrates:
VII. Conclusions
Source of Variation
Degrees of Freedom
Sum of Squares1
Mean Square
F2
Between Groups
2
101.47
50.74
0.47
Within Groups
41
4470.39
109.03
TOTAL
43
4571.86
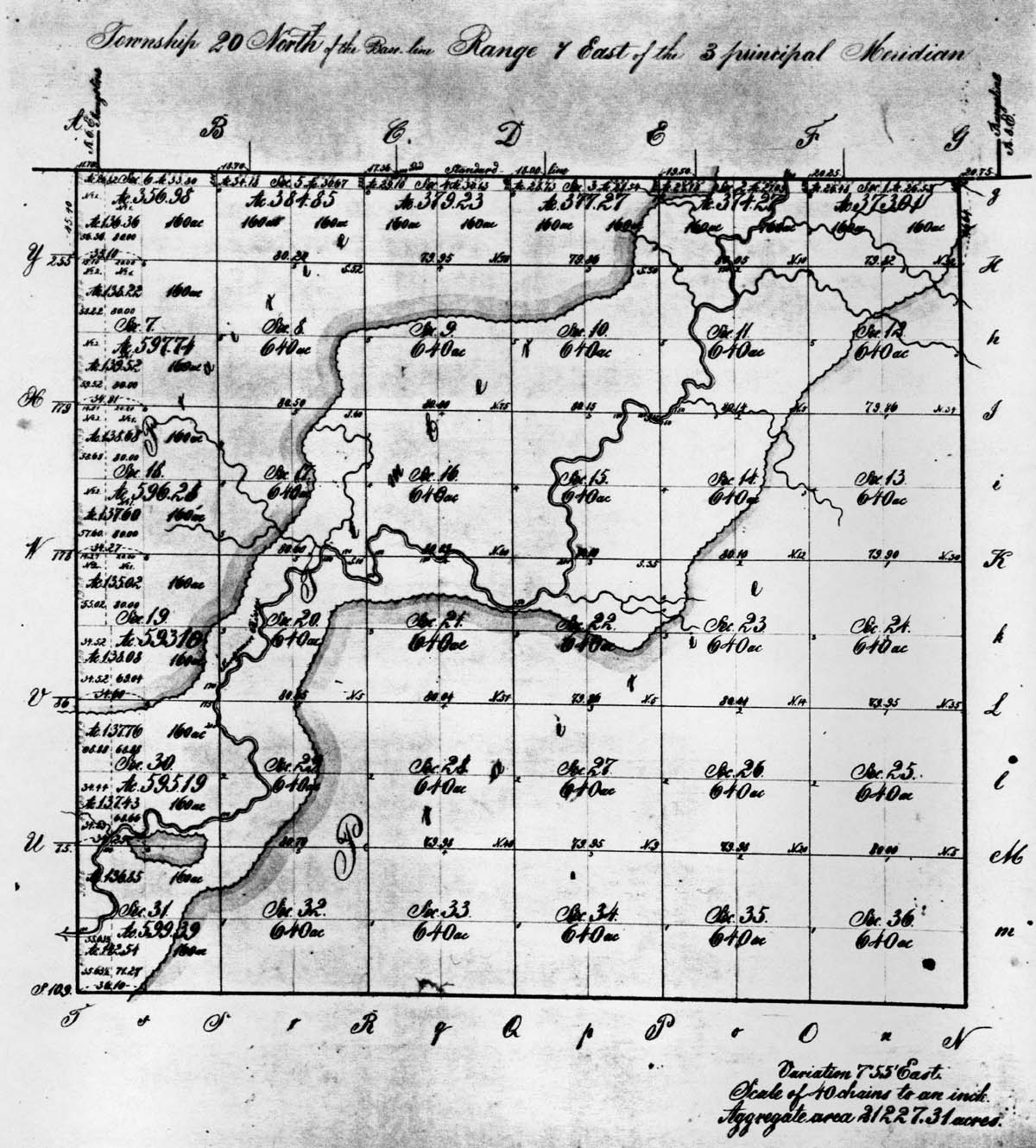
Table 2. Forest Species Identified in the GLO Land Survey (for a review
of the statistical measures employed in this analysis the reader is referred to
Cottam and Curtis [1956]; Rodgers and Anderson [1979]; and Boggess [1964]).
Species
Frequency
Relative Density
Relative Dominance
Importance Value
White Oak
31
58.5
48.8
107.3
Elm
6
11.3
10.9
22.2
Black Oak
5
9.4
11.1
20.5
Red Oak
3
5.7
8.0
13.6
Hickory
3
5.7
6.9
Bur Oak
1.9
7.0
8.9
Hackberry
1
1.9
4.8
6.7
Ash
1
1.9
1.2
3.1
Black Walnut
1
1.9
0.8
2.7
Willow
1
1.9
0.5
2.4
TOTALS
53
100.0
100.0
200.0
Total Distance = 400.9 m
Mean Distance = 400.9/33 = 12.1 m
Mean Area = (12.1)2 = 146.4 m2
Trees per ha = 10000/146.4 = 68.3
Avg Basal Area per Tree = 0.2 m2
Total Basal Area = 8.9 m2
Table 3. Importance Values (IV) for the First Five Dominants of the
Upland Forest in Five Studies of the Sangamon Drainage.
Species
Period/Site
Presettlement Mahomet Township
Presettlement Sangamon Basin1
Presettlement Pabst Site2
Historic Hart3
Historic Allerton4
Combined
Slope
Upland
Combined
Slope
Upland
Upland
White Oak
107.3
69.5
50.8
112.1
40.1
98.1
50.3
Black Oak
20.5
67.5
78.8
50.6
26.4
57.4
30.6
Elm
22.2
14.4
12.0
6.7
39.9
14.2
25.2
Hickory
12.6
26.0
41.7
22.7
--
--
36.0
Basswood
--
--
7.6
3.8
--
--
--
Black Walnut
--
4.1
--
--
--
--
--
Red Oak
13.6
--
--
--
31.8
11.9
12.3
Black Cherry
--
--
--
--
--
6.5
--
Shingle Oak
--
--
--
--
9.1
--
--
Table 4. Importance Value of Five Leading Dominants of the Floodplain
Forest From Three Sites in the Sangamon Drainage.
Species:
Period/Site
Presettlement Central Sangamon1
Historic Allerton2
Historic Hart3
Elm
44.6
21.6
36.6
Silver Maple
--
75.2
63.8
Sycamore
24.0
--
--
Hickory
19.0
--
--
Oak
46.4
37.5
--
Black Walnut
19.5
--
11.7
Green Ash
--
10.6
30.1
Hackberry
--
24.6
14.1
Table 5. Historically Reported Mammals, Preferred Habitats, and Presence in the Archaeological Record.
Species1:
Preferred Habitat:
Found in Archaeological Context2 :
Oppossum
Forest/wooded streams
Yes
Shorttail shrew
All terrestrial environments
Yes
Southeastern shrew
Open forest/forest edge
No
Least shrew
Forest edge/prairie
Yes
Eastern mole
All upland habitats
Yes
Star-nosed mole
All upland habitats
No
Little brown bat
Roosts in trees
No
Keen bat
Roosts in trees
No
Indiana bat
Roosts in trees
No
Small footed bat
Roosts in trees
No
Silver haired bat
Roosts in trees
No
Eastern pipistrel
Roosts in trees
No
Red bat
Roosts in trees
No
Big brown bat
Roosts in trees
No
Hoary bat
Roosts in trees
No
Evening bat
Roosts in trees
No
Raccoon
Wooded bluffs/floodplain
Yes
Marten
Dense forest
Yes
Fisher
Floodplain forest
Yes
Least weasel
Stream borders/dry prairie
No
Longtail weasel
Stream borders/dry prairie
No
Mink
Along all water courses
Yes
River otter
Sangamon
Yes
Badger
Dry to mesic prairie
Yes
Striped skunk
Open forest/forest edge
Yes
Black bear
Floodplain forest
Yes
Domestic dog
(see Note 3 below)
Yes
Timber wolf
Wooded bluffs/forest
Yes
Coyote
Forest edge/prairie
No
Red fox
Wooded bluffs/forest edge
Yes
Gray fox
Dense forest
Yes
Puma
Dense forest
No
Bobcat
Dense forest/wooded bluffs
Yes
Woodchuck
Wooded bluffs/forest
Yes
Thirteen-lined ground squirrel
Dry prairie
Yes
Franklin ground squirrel
Dry to mesic prairie/forest edge
No
Eastern chipmunk
Wooded bluffs/river bottoms
Yes
Eastern gray squirrel
Dense forest/river bottoms
Yes
Eastern fox squirrel
Open forest/forest edge
Yes
Red squirrel
Dense forest
Yes
Southern flying squirrel
Dense forest/wooded bluffs
No
Plains pocket gopher
Prairie
Yes
Beaver
Sangamon/wooded streams
Yes
Deer mouse
Prairie
Yes
White footed mouse
Wooded bluffs/floodplain
Yes
Golden mouse
Floodplain forest
Yes
Southern bog lemming
Open forest/floodplain
Yes
Meadow vole
Wet meadows/floodplain forest
Yes
Prairie vole
Prairie/forest edge
Yes
Pine vole
Forest
Yes
Muskrat
Sangamon/streams/prairie sloughs
Yes
Norway rat
Cultural disturbance
Introduced
Black rat
Cultural disturbance
Introduced
House mouse
Cultural disturbance
Introduced
Meadow jumping mouse
Wooded streams
Yes
Eastern Cottontail
All open habitats/forest edge
Yes
White tailed deer
Forest/forest edge/thickets
Yes
Elk
Open forest/wooded streams
Yes
Bison
Prairie/forest edge
Yes
1 - See Appendix for scientific names.
2 - Compiled from Lewis (1979), Parmalee and Klippel (1983),
Parmalee and Bogan (1981), and Purdue and Styles (1986).
3 - The reference here is to varieties associated with prehistoric
peoples.
Table 6. Birds Representative of Several Habitats within the Park.
Species Preferred Habitat
======== ==================
Natrix rhombifera - Diamondback water snake Aquatic
Masticophis flagellum - Whip snake Upland Prairie
Elaphe guttaga - Rat snake Upland Prairie
Pituophis catenifer - Bull snake Upland Prairie
Lampropeltis getulus - Speckled kingsnake Upland Forest
Ancistrodon contortrix - Copperhead Upland Forest
Crotalus horridus - Timber rattlesnake Forest
Kinosternon spp. - Mud turtle Aquatic/mud bottom
1 - Adapted from Zetek (1918:154).
Kingdom: Animalia
Phylum: Chordata
Subphylum: Vertebrata
Class: Mammalia
Subclass: Theria
Infraclass: Metatheria
Order: Marsupialia (Marsupials)
Family: Didelphiidae
Didelphis marsupials - Opposum
Infraclass: Eutheria
Order: Insectivora
Family: Soricidae (Shrews)
Sorex longirostris - Southeastern shrew
Cryptotis parva - Least shrew
Blarina brevicauda - Shorttail shrew
Family: Talpidae (Moles)
Scalopus aquaticus - Eastern mole
Condylura cristate - Star-nosed mole
Order: Chiroptera (Bats)
Family: Vestertilionidae
Myotis lucifugus - Little brown bat
Myotis keeni - Keen bat
Myotis sodalis - Indiana bat
Myotis subalatus - Small footed bat
Lasionycteris noctivagaus - Silver haired bat
Pipistrellus subflavus - Eastern pipistrel
Lasiurus borealis - Red bat
Eptesicus fuscus - Big brown bat
Lariurus cinereus - Hoary bat
Nycticeius humeralis - Evening bat
Order: Carnivora (Flesh-eaters)
Family: Procyonidae (Raccoons)
Procyon lotor - Raccoon
Family: Mustelidae (Weasels, Skunks)
Mustela americana _ Marten
Mustela canadensis - Fisher
Mustela rixosa - Least weasel
Mustela frenata - Longtail weasel
Mustela vison - Mink
Lustra canadensis - River otter
Taxidea taxus - Badger
Mephitis mephitis - Striped skunk
Family: Canidae (Wolves, Foxes)
Canis lupus - Timber wolf
Canis latrans - Coyote
Vulpes fulva - Red fox
Urocyon cinereoargenteus - Gray fox
Family: Ursidae (Bears)
Ursus americanus - Black bear
Family: Felidae (Cats)
Felis concolor - Puma
Lynx rufus - Bobcat
Order: Rodentia (Gnawing Mammals)
Family: Sciuridae (Squirrels)
Marmota monas - Woodchuck
Citellus tridecemlineatus - Thirteen-lined ground squirrel
Citellus franklini - Franklin ground squirrel
Tamias striatus - Eastern chipmunk
Sciurus carolinensis - Eastern gray squirrel
Sciurus niger - Eastern fox squirrel
Tamiasciurus hudsonicus - Red squirrel
Glaucomys volans - Southern flying squirrel
Family: Geomyidae (Pocket gophers)
Geomys bursarius - Plains pocket gopher
Family: Castoridae (Beaver)
Castor canadensis - Beaver
Family: Cricetidae (Mice, Lemmings and Voles)
Peromyscus maniculatus - Deer mouse
Peromyscus leucopus - White-footed mouse
Peromyscus nuttalli - Golden mouse
Synaptomys cooperi - Southern bog lemming
Microtus pennsylvanicus - Meadow vole
Microtus ochrogaster - Prairie vole
Pitymys pineotorum - Pine vole
Ondatra zibethicsa - Muskrat
Family: Muridae (Old Word Rats and Mice)
Rattus norvegicus - Norway rat
Rattus rattus - Black rat
Mus musculus - House mouse
Family: Zapodidae (Jumping Mice)
Zapus hudsonius - Meadow jumping mouse
Order: Lagomorpha (Pikas, Hares and Rabbits)
Family: Leporidae (Hares and Rabbits)
Sylvilagus floridanus - Eastern cottontail
Order: Artiodactyla (Even-Toed Hoofed Mammals)
Family: Cervidae (Deer)
Odocoileus virginianus - White-tail deer
Cervus canadensis - Elk
Family: Bovidae (Bison)
Bison bison - Bison
Order: Primates
Suborder: Anthropoidea
Superfamily: Hominoidea
Family: Hominidea
Genus: Homo
Species: sapiens
Variety: sapiens